A high-resolution map of recombination in maize.
Results
I. Generating a map of sites in the maize genome where recombination is initiated by formation of meiotic double-strand-breaks (DSBs) in chromosomal DNA.
- Using a chromatin immunoprecipitation approach (ChIP), we generated a map of meiotic DSB hotspots in the B73 maize inbred, which is the first high-resolution map of recombination hotspots in any plant species.
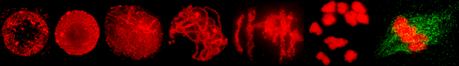
- To validate our DSB hotspot map, we examined DSB formation at centromeres. Maize DSB hotspots are fairly evenly distributed along chromosomes, including the centromeres. However, centromeric regions are known to be devoid of COs and in yeast and mammals have also been shown to lack DSBs. To confirm the existence of cetromeric DSBs in maize, we conducted experiments combining the anti-RAD51 antibody with a fluorescence in situ hybridization (FISH) probe detecting the maize centromere repeat. We found that indeed RAD51 foci were present at several centromers in mid-zygotene meiocytes.
- To examine whether hotspot sites vary depending on the genetic background, we generated DSB maps in the Mo17 inbred and the B73 x Mo17 hybrid. We found that genomic features of DSB hotspot regions were conserved, including presence of a positive correlation with GC content, open chromatin and the consensus sequence motif. However, while some hotspots in Mo17 were at the same positions as in B73, we also found novel hotspots absent from B73. In the B73 x Mo17 hybrid, we also found novel hotspots absent from B73 and Mo17. These observations imply that locations of recombination hotspots are affected by trans-acting as well as cis-acting factors. We are now generating DSB maps for CML228 and the B73 x CML228 hybrid, which will be completed in summer of 2015.
- We examined DSB dynamics in heterochromatin using co-immunocolocalization of RAD51 with a heterochromatin marker H3 histone lysine 9 dimethylation (H3K9me2). We observed strong H3K9me2 staining throughout prophase I. As the genome of maize is heavily heterochromatic, the staining was too ubiquitous to allow counting of individual H3K9me2 foci. Instead, we delineated in three-dimensional microscopic images domains of H3K9me2 signal, discernable as areas containing large numbers of H3K9me2 foci on chromatin. Then, we quantified RAD51 foci in relation to the H3K9me2 domains. We found that when RAD51 foci first appear on chromosomes in leptotene, only about a quarter of them occur within H3K9me2 domains. However, later on, during zygotene, as many as 60% of RAD51 foci colocalized with H3K9me2-marked heterochromatin. These observations indicate that when the first DSBs are formed during maize meiosis, most of them are in euchromatin, However, later on, most DSBs are formed in heterochromatin. It has been proposed that a DSB fate may depend on the timing of its formation; DSBs made earlier are more likely to be repaired as COs than those made later. This mechanism might be behind the fact in maize few CO are formed in heterochromatin even though most DSBs are present in heterochromatic regions.
.
II. Mapping at high-resolution the sites where recombination events are resolved into reciprocal chromosome exchanges (crossovers, COs).
To generate CO maps in maize, we used a genetic approach, which at present is the only reliable way of CO mapping. We have now completed genotyping two backcross populations: B73 X (B37 X Mo17) to examine recombination in male meiosis in the B73 x Mo17 hybrid and (B37 X Mo17) X B73, which is a reciprocal population set up to examine recombination in female meiosis. Together, from these two populations we sequenced 291 individuals at an average coverage of ca. 1.9 x. The average marker spacing in these populations is about 2kb. However in genic regions, which contain most COs, the spacing goes down to an average of about 600bp. These two populations are being analyzed to determine the overall patterns of CO distribution, the relationship between CO distribution in male vs. female meiosis, and the association of CO sites with DNA sequence and chromatin features.
We found that there are no statistically significant differences between the number of COs per meiosis in males vs. females in the B73 x Mo17 cross. However, there are substantial differences in how COs are distributed along chromosomes. While distal regions of chromosomes showed similar patterns in male and female meiosis, chromosomes 1, 4, and 10 showed higher CO rates in pericentromeric regions in female meiosis and chromosomes 8 showed higher CO rates in the pericentromeric region in male meiosis.
We found substantial differences in CO distribution in male vs. female meiosis even in regions where the overall CO rates in males and females were similar, indicating that locations of recombination hotspots differ between male and female meiosis.
.
III. Relating the maps of DSB and CO distribution to local chromatin features.
We examined the transcriptome of isolated leptotene and zygotene meiocytes in B73, Mo17, and CML228.
We mapped nucleosome occupancy in leptotene-zygotene anthers of B73.
We mapped H3K4 tri-methylation sites in isolated meiocytes in B73, Mo17, and CML228.
We examined meiosis-specific small RNAs in B73 and Mo17.
We analyzed meiosis-specific DNA methylation patterns in B73.
Copyright © 2015 Cornell University